Пересказ статьи:
Mahdokht Jouiaei, Angel A. Yanagihara, Bruno Madio, Timo J. Nevalainen, Paul F. Alewood, Bryan G. Fry. 2015
Ancient Venom Systems: A Review on Cnidaria Toxins
Toxins. 7(6), 2251-2271
https://doi.org/10.3390/toxins7062251
Обзорная статья, 146 литературных источников. Слегка приглаженный машинный перевод. Часть текста заменена ссылками на статьи этого сайта. Перевод биохимических терминов не проверялся специалистом.
Из приблизительно 10 000 видов Cnidarian было выделено 156 токсинов 10 фармакологических семейств. Потенциально полезная новая токсикологическая информация постепенно накапливается. Одним из многообещающих решений для расширения знаний о токсинах Cnidarian является применение высокопроизводительных технологий секвенирования для выявления новых структурных и фармакологических групп токсинов.
Огромный потенциал ядов Cnidarian для понимания активности различных рецепторов и каналов в состоянии здоровья и заболеваний, а также их использования в качестве источников новых фармацевтических соединений открывает бесконечные возможности для будущих научных исследований.
1. Введение
Характеристика типа. Можно читать у меня
2. Филогения книдарий
На основании данных митохондриальной ДНК (мтДНК) и жизненных циклов книдарии делятся на две ныне живущие подгруппы: Anthozoa и Medusozoa. Anthozoa обладают кольцевой мтДНК, сходной с таковой у других многоклеточных животных, в то время как у Medusozoa есть нетипичная линейная мтДНК. Представители относящихся к Medusozoa классов Hydrozoa, Schyphozoa, Cubozoa и Staurozoa демонстрируют трехфазный жизненный цикл с переходом поколений: стадия свободноплавающей личинки «планула», стадия сидячего полипа и стадия половозрелой пелагической медузы. У Anthozoa стадия медузы потеряна, и сидячие взрослые особи осуществляют половое размножение. Жизненный цикл будет обсуждаться более подробно далее в этом обзоре.
3. Жизненный цикл книдарий
Можно читать у меня
4. Система доставки яда Cnidaria
Можно читать у меня
5. Состав яда
С начала 20-го века многочисленные аналитические исследования и клинические наблюдения установили токсикологическое разнообразие ядов стрекающих (Таблица 1). Разнообразие компонентов яда варьируется от небелковых соединений (например, пуринов, биогенных аминов) до высокомолекулярных белков, эволюционировавших в течение сотен миллионов лет. Интересно, что некоторые типы семейств токсинов, идентифицированные ранее у других ядовитых животных, составляют арсенал яда у кишечников. Наиболее ярким примером является то, что пептиды Кунитц-типа ясно вырожены у актиний, улиток-конусов, насекомых, скорпионов, пауков, клещей, рептилий , и летучих мышах-вампирах. Другим примером конвергентной экспрессии токсинов является блокатор калиевых каналов Kv1, который не только конвергентно развился у скорпионов и актиний, но даже использует одни и те же остатки для блокирования пор каналов.
В оригинальной таблице есть ссылки на источники.

5.1 .Ферменты
5.1.1. Фосфолипаза А2
Активность фосфолипазы А2 (PLA2) была обнаружена в гомогенизированных щупальцах и аконтиях книдарий, включая подклассы Anthozoa, Schyphozoa, Hydrozoa и Cubozoa. Этот фермент гидролизует sn-2 ацильную связь глицерофосфолипидов с образованием жирных кислот, включая арахидоновую кислоту и лизофосфолипиды. Он обычно обнаруживается в тканях млекопитающих, где он играет важную роль в жизненно важных функциях организма, включая катаболизм липидов в рационе, воспаление, передачу сигнала и ремоделирование фосфолипидов. Тем не менее, повсеместное присутствие PLA2 в ядовитых группах животных, таких как рептилии (змеи и ящерицы надотряда Anguimorpha), многоножки, насекомые (их щетина, хоботки и жала), паукообразные (скорпионы, пауки и клещи), книдарий и головоногие указывает на конвергентный набор белков организма в токсичный арсенал животных. Предполагается, что токсические функции PLA2 в ядах книдарий включают защиту, иммобилизацию и переваривание добычи. Также гемолитическая активность была продемонстрирована во фракции PLA2, выделенной из аконистических нематоцист актинии Aiptasia pallida. Несмотря на то, что энзимы PLA2 многочисленны среди книдарий, связь их молекулярной структуры и функций еще не выяснена.
5.2. Порообразующие токсины
По-видимому, порообразующие токсины (PFT) присутствуют во всех ядах книдарий. Механизм действия этих токсинов заключается в проникновении через клеточную мембрану мишени, что приводит к диффузии малых молекул и растворенных веществ, что приводит к осмотическому дисбалансу и лизису клеток. PFT обладают двойственной структурой: (i) стабильная водорастворимая структура, которая является мономерной и связывается с рецепторами на клетке-мишени; (ii) мембраносвязанная структура, состоящая из олигомерных молекул, которые образуют цельные мембранные поры. PFT книдарий подразделяются на две группы в зависимости от типа вторичной структуры, которую они используют для проникновения через мембрану при порообразовании: α-PFT, где содержится большое количество спиралей и образуются α-спиральные цилиндрические структуры, и β-PFT, которые богаты β-пластинами и образуют β-цилиндрические поры.
5.2.1. Актинпорины
Актинпорины представляют собой α-PFT и присутствуют у Anthozoa и Hydrozoa. Они представляют собой основные белки размером примерно 20 кДа, в которых отсутствуют внутримолекулярные дисульфидные связи. Актинопорины опосредуют различные виды токсичности и биоактивности, вызванные порообразующими механизмами, такие как остановка сердечно-сосудистой системы и дыхания у крыс, лизис эритроцитов кур, коз, человека и овец и цитотоксические эффекты. В нескольких исследованиях сообщалось, что актинопорины взаимодействуют исключительно с сфингомиелинсодержащими мембранами, хотя стихолизин II из Stichodactyla helianthus связывается с фосфатидилхолиновой мембраной. Механизм проникновения через мембрану состоит из нескольких этапов: (i) первоначальное связывание с мембраной-мишенью, которое осуществляется открытой петлей, богатой ароматическими веществами; (ii) боковая ориентация и последующая олигомеризация с помощью сайта связывания фосфоколина; и (iii) вставка N-концевого амфифильного α-спирального сегмента в липидную мембрану. Ядерный магнитный резонанс высокого разрешения и эксперименты по инфракрасной спектроскопии с Фурье-преобразованием, измерения собственной флуоресценции, рентгеновская кристаллография и электронно-микроскопический анализ двумерных кристаллов показали, что во время проникновения через мембрану N- концевой амфифильный α-спиральный сегмент актинопоринов отделяется от основного тела и вставляется в мембрану-мишень с образованием пор диаметром 1–3 нм (~ 11–30 Å)
5.2.2. Токсины медуз (JFTs)
Порины кубомедуз Cubozoa являются наиболее сильнодействующими и быстродействующими токсинами, из выделяемыми разными медузами. Это семейство токсинов было первоначально зарегистрировано у кубомедузы Catabdea alata как CAH1, также известный как CaTX-A / B, а затем идентифицировано у всех исследованных кубомедуз, включая CrTX-A / B из Carybdea rastoni, CqTX -A из Chiropsalmus quadrigatus , CfTX-1/2 и CfTX-A / B / Bt из Chironex fleckeri. Тем не менее, гомологи поринов кубомедуз были обнаружены у Scyphozoa (Aurelia aurita), Hydrozoa (Hydra magnipapillata), Anthozoa (Aiptasia pallida) и различных гидроидов (Hydractinia symbiolongicarpus и Hydra vulgaris), что указывает на общее эволюционное происхождение этих ядов. Характерно, что они являются основными белками с молекулярной массой от 40 до 46 кДа и содержат как α, так и β домены. Гипотетические механизмы, лежащие в основе образования пор, включают олигомеризацию нескольких амфифильных и гидрофобных α-спиралей в N-концевой области токсина, что приводит к повреждению плазматической мембраны и гибели клеток. Сходство в трехмерной структуре N-концевого домена токсинов CfTX со структурой α-порообразующего домена (домен I) инсектицидных δ-эндотоксинов из Bacillus thuringiensis позволяет предположить, как токсины этого семейства внедряются в мембраны. Этот механизм мембраных вставок, по-видимому, достаточен для объяснения четко определенных пор диаметром 12 нм (внутренний) и 25 нм (внешний), создаваемых в эритроцитах человека токсинами CfTX.
Интересно, что члены этого семейства токсинов демонстрируют различную специфичность к различным тканям позвоночных. Недавнее исследование токсинов CfTX показало, что CfTX-1/2 вызывает сердечно-сосудистый коллапс в течение 1 мин у анестезированных крыс, подвергшихся действию яда in vivo, тогда как CfTX-A / B более активен в проявлении гемолитической активности in vitro. Хотя эти данные свидетельствуют о том, что семейство токсинов медуз претерпело функциональную диверсификацию, необходимы дополнительные структурные / функциональные данные, чтобы понять истории эволюции молекул, которые сформировали эту диверсификацию.
5.2.3. Токсины, связанные с гидролизинами (Hydralysins)
В дополнение к поринам, обнаруженным у книдарий, в последних исследованиях описано новое семейство β-PFT, секретируемое из пищеварительных энтодермальных клеток зеленой гидры (Chlorohydra viridissima). Предполагается, что эти не-нематоцистные токсины, производимые организмом, выделяемые во время питания, играют роль в лизировании тканей жертвы. Эти токсины не активны в отношении мембранных фосфолипидов или углеводов, а скорее связываются со специфическими мембранными рецепторами и образуют поры. Недавнее исследование охарактеризовало аэролизин, гомолог гидролизина, секретируемый из эктодермальной клетки глотки актинии Nematostella vectensis, и предположило его роль в переработке добычи.
5.2.4. Комплекс мембранной атаки — перфорин
Эта группа β-PFT токсинов была обнаружена в ядах, извлеченных из актиний Phyllodiscus semoni и Actinaria villosa. Токсины могут быть использованы при захвате добычи / защите от хищников. Белки мембранного атакующего комплекса (MAC) были идентифицированы в системе комплемента, продуцируемой Т-клетками и NK-клетками. Белки создают трансмембранную пору в клетке-мишени и инициируют различные пути гибели апоптотических клеток.
5.3. Нейротоксины
Нейротоксины книдарий (потенциал-зависимые токсины ионных каналов) представляют собой группу низкомолекулярных пептидов и являются одними из наиболее изученных токсинов с точки зрения механизма действия. Они производятся актиниями и играют фундаментальную роль в их яде, помогая этим сидячим животным быстро обездвиживать добычу и защищаться от хищников. Они продлевают потенциал действия возбудимых и невозбудимых мембран в сенсорных нейронах и клетках сердечной и скелетных мышц путем модификации шлюзов натриевых каналов или блокирования шлюзов калиевых каналов на стадии реполяризации. Это приводит к тому, что клетка становится гиперактивной и высвобождает огромное количество нейротрансмиттеров в синапсах и нервно-мышечных соединениях, что может вызвать первоначальную спастическую стадию с последующим нисходящим вялым параличом. Токсины шлюзов ионных каналов актиний были тщательно изучены, поскольку они являются ценными биоресурсами для изучения структуры и функции натриевых и калиевых каналов, а также для разработки лекарственных средств и биоинсектицидов.
Некоторые полипептиды нейротоксинов Cnidarian являются исключительно блоком ванилойдных рецепторов 1 (TRPV1) и чувствительным к кислоте ионным каналом 3 (ASIC3), которые участвуют в инициации и передачи боли и гипералгезии. Таким образом, эти пептиды являются многообещающими инструментами для разработки новых болеутоляющих средств.
5.3.1. Токсины натриевых каналов
Потенциал-зависимые токсины натриевых (Nav) каналов, (NaTxs), представляют собой трансмембранные комплексы, состоящие из нескольких субъединиц. Высококонсервативная α-субъединица состоит из четырех гомологичных доменов (D1 – D4), каждый из которых содержит шесть гидрофобных трансмембранных областей (S1 – S6). Anthozoan NaTxs и некоторые другие группы токсинов из скорпиона и пауков связываются с сайтом 3 (петля S3-S4 в D4) каналов Nav, что приводит к значительному выбросу нейротрансмиттера в синапсах. Одним из возможных объяснений его токсичности является электростатическое взаимодействие между кластером основных аминокислот на токсине и кислотными аминокислотами в сайте 3, блокирующее сегмент S4 во его внутреннем положении, что препятствует конформационным изменениям канала, необходимым для быстрой инактивации. Эти токсины делятся на три группы: (i) NaTx типа I; и (ii) NaTxs типа II обладают обширным сходством последовательностей и имеют сходную функцию. В дополнение к NaTxs типа I и II существует сиротливая клада NaTx, которая проявляет только частичную гомологию как к NaTxs типа I, так и типа II и демонстрирует механизм действия, сходный с таковым. Наконец, в яде актиний присутствуют короткие (длиной ~ 30 аминокислот) пептиды, которые проявляют активность, аналогичную остальным семействам NaTxs, несмотря на отсутствие каких-либо общих мотивов последовательности с остальными из них. Эти пептиды образуют семейство типа III и проявляют очень высокую селективность по отношению к натриевым каналам членистоногих.
5.3.2. Потенциал-зависимые токсины калиевых каналов (Kv) и пептиды Кунитца.
Токсины потенциал-заисимых калиевых каналов (Ktxs) актний подразделяются на пять групп по сходству последовательностей и сродству связывания с различными семействами Kv каналов. KTx типа I имеют молекулярную массу 4 кДа с тремя дисульфидными мостиками. Они подавляют ток калия по подсемейству каналов Kv1. и Kv3 и промежуточную проводимость кальций-активированных калиевых каналов. KTx типа II представляют собой пептиды Кунитца с молекулярной массой 6 кДа, которые сшиты тремя дисульфидными мостиками и интересны двойственностью функций. Обычно они действуют против трипсиновых и химотрипсиновых протеиназ, чтобы ингибировать быструю деградацию ядовитой протеазы эндогенными ферментами самих животных или жертвы. Интересно, что некоторые пептиды Kуница актиний обладают активностью блокирования Kv, сходной с активностью ингибирования дендротоксина и протеазы. Например, каликлудин 1-3 из A. sulcata конкурентно связывается с каналами Kv1.2, чтобы быстро парализовать добычу. KTx типа III представляют собой пептиды 3-4 кДа с тремя дисульфидными мостиками, которые развились из NaTxs в режиме положительного отбора. Они блокируют множество различных каналов ионов калия, таких как канал Kv3.4, принадлежащих к быстро инактивирующемуся каналу Kv и hERG (человеческий Ether-à-go-go-Related ген). Интересно отметить, что влияние положительного отбора не приводит к полной потере активности блокирования ионно-натриевых каналов, когда несколько KTx типа III нацелены как на hERG, так и на чувствительный к кислоте ионный канал 3 (ASIC3, H + -зависимые Nav каналы). Тип IV — структурно новые пептиды актинии Stichodactyla haddoni, которые парализуют крабы, сшитые двумя дисульфидными мостиками. Тип V был обнаружен у актинии Bunodosoma caissarum и сшит четырьмя дисульфидными мостиками. Этот новый пептид активен на ИК-каналах Drosophila Shaker.
5.3.3. Маленькие пептиды, богатые цистеином (SCRiPs)
Эта недавно обнаруженная группа нейротоксинов была обнаружена в эктодерме рифообразующих кораллов Acropora millepora и гомологи SCRiP были обнаружены у актиний Anemonia viridis и Metridium senile. Инъекция рекомбинантно экспрессированных SCRiP A. millepora личинкам рыбок данио (Danio rerio) привела к тяжелому параличу, что указывает на то, что это первое семейство пептидных нейротоксинов, описанное у склерактиновых кораллов.
5.3.4. Ингибиторы ASIC
ASIC — это натрий-селективные кислоточувствительные ионные каналы (ASIC), экспрессируемые в периферической нейронной системе. Они связаны с кислотной болью во время патологических состояний, таких как воспаление и ишемия. Недавно сообщалось, что новый пептид π-AnmTX Ugr 9a-1 из яда актинии Urticina grebelnyi и PhcrTx1 из Phymanthus crucifer имеют своими мишенями каналы ASIC. Эти пептиды сшиты двумя дисульфидными мостиками и не имеют последовательности гомологичной другими пептидами нейротоксинов актиний.
5.3.5. Ингибиторы TRPV1
TRPV1 представляют собой неселективные катионные каналы, экспрессируемые в периферической и центральной нейронных системах млекопитающих. Они инициируют нейронный ответ на воспалительные стимулы, что позволяет рассматривать их как один из наиболее важных молекулярных триггеров болевых стимулов. Первым пептидным ингибитором TRPV1, выделенным из яда актиний был τ-SHTX-Hcr2b (APHC1) из Heteractis crispa. Впоследствии два гомологичных пептида (τ-SHTX-Hcr2c (APHC2) и τ-SHTX-Hcr2d (APHC3), которые нацелены на TRPV1, были выделены из этого же вида. Эти новые нейротоксины являются многообещающими новыми моделями для разработки анальгетиков нового поколения.
5.4. Небелковые биоактивные компоненты
В дополнение к белковым и пептидным соединениям, упомянутым выше, в ядах книдарий был обнаружен ряд фармакологически активных низкомолекулярных соединений. Сообщалось о большом количестве 5-гидрокситриптамина (5-НТ, серотонина) в воде, где гидру стимулировали электрически. Вероятно, это производное нематоцист, которое вызывает мгновенную боль у хищников и, таким образом, может играть защитную роль. Расширение кровеносных сосудов, усиленое серотонином, может усиливать действие других компонентов яда. Гистамин был обнаружен в гомогенизированных щупальцах актиний Anemonia viridis (ранее известный как Anemonia sulcata) и Actinia equina. Как и серотонин, гистамин вызывает сильную боль и увеличивает проницаемость сосудов. Хотя эти соединения не были непосредственно обнаружены в изолированных нематоцистах, вполне вероятно, что они причастны к ядовитости книдарий, и для облегчения симптомов было рекомендовано лечение поражений ядом антигистаминными препаратами. Бунодозин, N-ациламинокислота, был выделен из яда актинии Bunodosoma cangicum. Это соединение проявляет сильную анальгетическую активность при активации рецепторов серотонина. Кессарон представляет собой производное четвертичного пурина, выделенное из морского анемона Bunodosoma caissarum путем экстракции всего животного в ацетоне. Этот морской продукт обладает высокой антагонистической активностью в отношении аденозиновых рецепторов подвздошной кишки морской свинки, которые стимулируют функцию кишечника.
6. Роль яда Cnidarians в разработке лекарств
(пропущено при переводе)
7. Интоксикация от ядовитого укуса
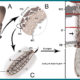
К счастью, из-за биофизических свойств разряд стрекательных клеток многих книдарий не способны проникать в кожу человека. А те укусы большинства видов книдарий, которые способны проникать в ткани человека, приводят лишь к незначительному или умеренному кратковременному раздражению/жжению. Но вот контакт с несколькими видами медуз, актиний и кораллов может вызвать сильную боль, повреждение тканей и даже сердечно-сосудистый коллапс, приводящий к смерти. Яд опасных животных вводится в ткань (рис. 1) и инициирует немедленные и отсроченные иммунологические и токсикологические реакции. Симптомы отравления определяются содержанием яда, объемом инъекции, состоянием здоровья пациента и продолжительностью контакта щупальца с кожей.
Одним из наиболее важных шагов в лечении поражений человека является использование специфических жидкостей для предотвращения дальнейшего выделения нематоцист, поскольку физические попытки оторвать оставшиеся щупальца от кожи жертвы могут вызвать массивные выстрелы нематоцист. С 1908 года многочисленные традиционные вещества, такие как мочевина, морская вода, уксус, метилированные спирты и т. д использовались для инактивации неизрасходованных нематоцист на прилипших щупальцах и / или для облегчения боли. Эти химические вещества были использованы с различной степенью успеха, начиная от полного успеха у одних до полного отказа у других. Например, в то время как некоторые химические вещества, такие как этанол или 5% уксусная кислота в дистиллированной воде, вызывают массивные срабатывание нематоцист у видов Hydrozoa и Cubozoa, было обнаружено, что некоторые другие, такие как пищевой уксус, инактивируют проникающие нематоцисты морской осы (кубомедузы Chironex fleckeri) быстро и полностью.
Профилактика и лечение людей, пораженных ядом книдарий становится все более важной задачей научного сообщества, требующей совместных усилий представителей медицинских, фармакологических, токсикологических и биологических дисциплин для развития лучшего понимания полевой экологии , а также эффективных терапевтических средств для минимизации патогенного воздействия на купальщиков. Это также может улучшить наше понимание пищевой сети и динамики этих древних и интригующих животных в экосистемах.